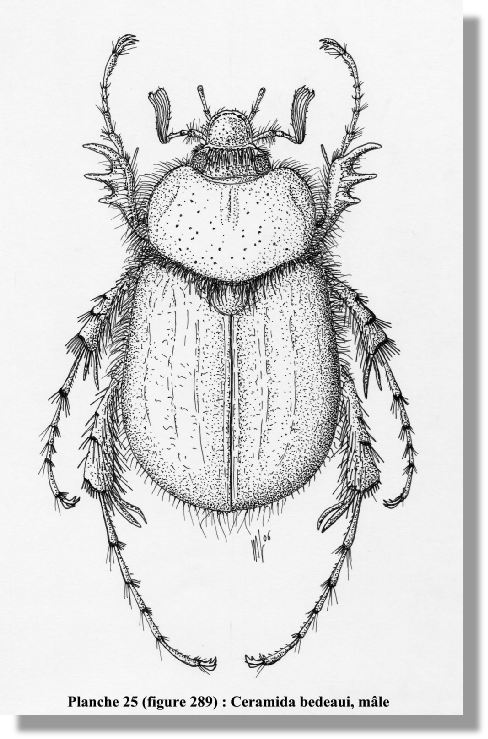
Description. – Corps assez massif, arrondi, sans revêtement pruineux au dessus (fig. 289). Dessus brillant, non pileux, d’un brun rougeâtre. Avant-corps plus foncé.
Clypéus important, allongé, à bords fortement relevés, formant cuvette ; bord antérieur bien arrondi (fig. 291). Ponctuation de la tête forte. Sillon clypéo-frontal marqué. Pilosité frontale le plus souvent longue, dense et dressée (fig. 298). Canthus oculaire présent. Yeux forts. Labre petit, peu visible, aplati (fig. 293). Dernier article des palpes maxillaires fortement allongé, affiné, cylindrique (fig. 292). Antennes de 10 articles, le III allongé, légèrement dilaté en son milieu. Massue antennaire de 7 articles chez le mâle (fig. 294).
Pronotum transverse, à côtés assez arrondis et longuement ciliés ; angles antérieurs arrondis ou peu marqués ; marge antérieure rebordée, avec une bande membraneuse (fig. 302). Disque non cilié, à ponctuation fine. Ecusson peu ponctué, non cilié.
Elytres à côtés élargis avant l’apex. Suture rebordée ; stries élytrales non marquées ou faibles. Calus huméraux et apicaux non marqués. Disque à ponctuation fine, non cilié. Apex légèrement rentré. Pygidium plus long que large, avec une longue ciliation latérale. Métasternum fortement villeux. Ventrites à ciliation éparse.
Protibia court, tridenté ; la dent basale bien marquée, la médiane longue, l’apicale longue et courbée (fig. 295). Eperon interne du protibia long, inséré en avant de la dent médiane. Tarses allongés. Articles II à IV des pro- et mésotarse fins, non élargis, sans brosses de soies en dessous. Méso- et métatibia sans carène transverse interne complète. Métafémur allongé. Métatibia court, élargi à l’apex (fig. 300) ; éperons terminaux longs, non dilatés, subégaux (fig. 299). Article I du métatarse bien plus long que le II (fig. 301). Griffes des tarses allongées, incisées à l’apex (fig. 296).
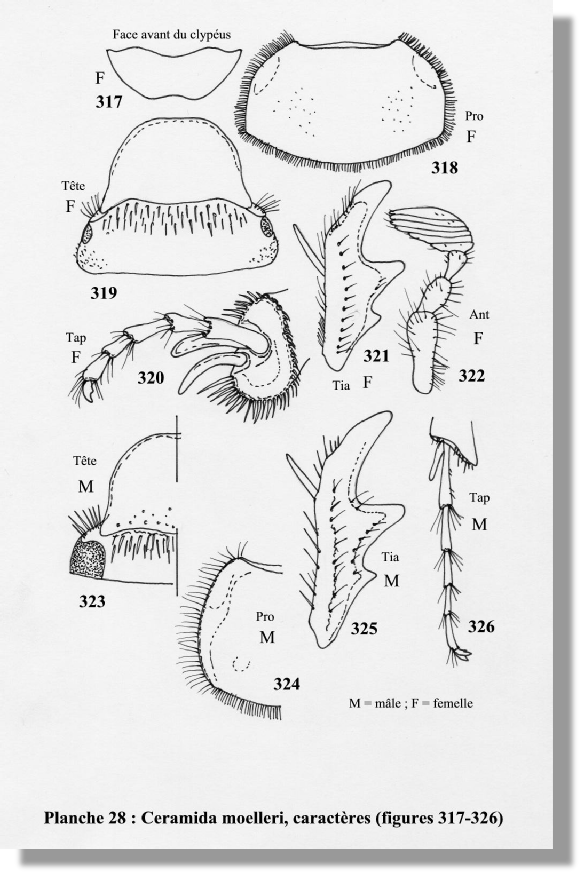
Femelle : Antennes avec parfois 9 articles dont 6 à la massue. Aptère, trapue, obèse. Yeux réduits. Pattes courtes. Protibia épaissi. Griffes des tarses courtes, réduites, non incisées, avec un denticule interne. Métafémur robuste. Métatibia fort. Métatarse court. Les femelles de longitarsis, malacensis et mauritanica n’ont pas été décrites ou observées.


Ceramida Baraud, 1987a : 126.
Ceramida Baraud ; Baraud, 1992 : 546.
Ceramida Baraud ; Sanmartín & al., 1998 : 26.
Ceramida Baraud ; Král & Smetana, 2006 : 199.
Ceramida Baraud ; Lacroix, 2007 : 49.
Elaphocera Géné ; Rambur, 1843 : 333.
Elaphocera Reitter, 1902 (nec Gené, 1836) : 97.
Sous-genre Elaphocera Reitt. ; Dalla Torre, 1913 : 305.
Elaphocera s. str. Reitter ; Báguena, 1955 : 123.
Elaphocera Gené s. str. ; Báguena, 1967 : 330, 331.
Elaphocera Gené (s . str.) ; Baraud, 1975 : 57.
Elaphocera Gené ; Baraud, 1977 : 272.
Elaphocera Gené ; Baraud, 1979a : 304.
Sous-genre Elaphocera ; Branco, 1981 : 125.
Elaphocera Géné (sic) ; Baraud, 1985 : 388.
Elaphocera Gené ; Serrano, 1985 : 360.
Elaphocera sensu Reitter, 1902, nec Gené, 1836 ; Baraud, 1987a : 126 (synonyme de Ceramida).
Elaphocera sensu Reitter, 1902 (non Gené) ; Sanmartín & al., 1998 : 26.
Espèce type : Melolontha longitarsis Illiger, 1803 (désignation par Baraud, 1987).
11 espèces

Biologie. – Voir : Rambur (1843) : bedeaui ; Cobos (1951) : bolivari, malacensis ; Branco (1981) : lusitanica ; Serrano (1985) : bedeaui ; Lopez-Colon (1993) : abderramani ; Sanmartin & Martin-Piera (1997) : cobosi.
Imago diurne. Espèces automnales. Mâles volant à la recherche des femelles et ne s’alimentant pas. Femelle enterrée près de la plante nourricière, laissant dépasser son abdomen. Vol des mâles le plus souvent pendant ou après de fortes pluies d’automne. Espèces côtières (Sud de la Péninsule ibérique). L’espèce mauritanica d’Afrique du Nord paraît avoir un cycle un peu différent (Lucas, 1849).
Discussion. – Le genre Ceramida Baraud comprend actuellement 11 espèces (Sanmartin & Martin-Piera, 1999b : 573-592) plus Ceramida ohausi (Reitter) sans doute récoltée en Espagne. Le nombre de taxons décrits s’élève à 29 (pour 11 bonnes espèces), huit espèces ayant été mises en synonymie en 1999 par les auteurs précités. Ceramida luisae (synonyme de C. baraudi Branco) est considéré par Arias & Lopez-Colon (2002) comme bonne espèce.
Ce genre, difficile d’étude (ainsi que Elaphocera), présente une grande confusion nomenclaturale avec la profusion d’espèces valides ou non selon les critères des auteurs, les combinaisons entre Elaphocera et Ceramida, les mises en synonymie nombreuses et quelquefois contradictoires (ex : huit taxons sous le nom de Ceramida malacensis). Le genre Ceramida est présent avant tout en Péninsule ibérique avec une espèce atteignant le Maroc et une endémique d’Afrique du Nord. Les femelles sont toujours très rarement récoltées.

|
Pays |
Portugal |
Espagne |
Maroc |
Algérie |
|
Espèces |
8 |
7 |
2 |
1 |
|
endémiques |
4 |
3 |
1 |
|

Liste des taxons décrits et correspondances
abderramani Escalera, 1923 > adusta Kraatz
adusta Kraatz, 1882 (variété de E. Bedeaui) > bonne espèce
baraudi Branco, 1981 > bonne espèce
bedeaui Erichson, 1840 > bonne espèce
bedeaui var. tangeriana Kraatz, 1882 > malacensis Rambur
bolivari Escalera, 1914 > mauritanica Rambur
brancoi Baraud, 1975 > bonne espèce
brandeiroi Flach, 1906 > bonne espèce
certa Báguena, 1955 > adusta Kraatz
championi Reitter, 1902 > bedeaui Erichson
cobosi Báguena, 1955 > bonne espèce
dinizi Branco, 1981 > bonne espèce
gigas Serrano, 1985 > moelleri Flach
isabellae Lopez-Colon, 1993 > moelleri Flach
longicornis Serrano, 1985 > brandeiroi Flach
longitarsis Illiger, 1803 > bonne espèce
luctuosa Báguena, 1955 > malacensis Rambur
luisae López-Colón & Rodriguez Arias, 1990 > baraudi Branco
lusitanica Baraud, 1975 > malacensis Rambur
malacensis Rambur, 1843 > bonne espèce
mancinii Báguena, 1955 > malacensis Rambur
mauritanica Rambur, 1843 > bonne espèce
moelleri Flach, 1906 > bonne espèce
numidica Rambur, 1843 > mauritanica Rambur
ohausi Reitter, 1902 > bonne espèce ?
onubensis Escalera, 1923 > bedeaui Erichson
tangeriana Kraatz, 1882 > malacensis Rambur
transtagana Branco, 1981 > baraudi, Branco
stenostoma Báguena, 1955 > bedeaui Erichson

Clé de détermination des espèces du genre (mâles)
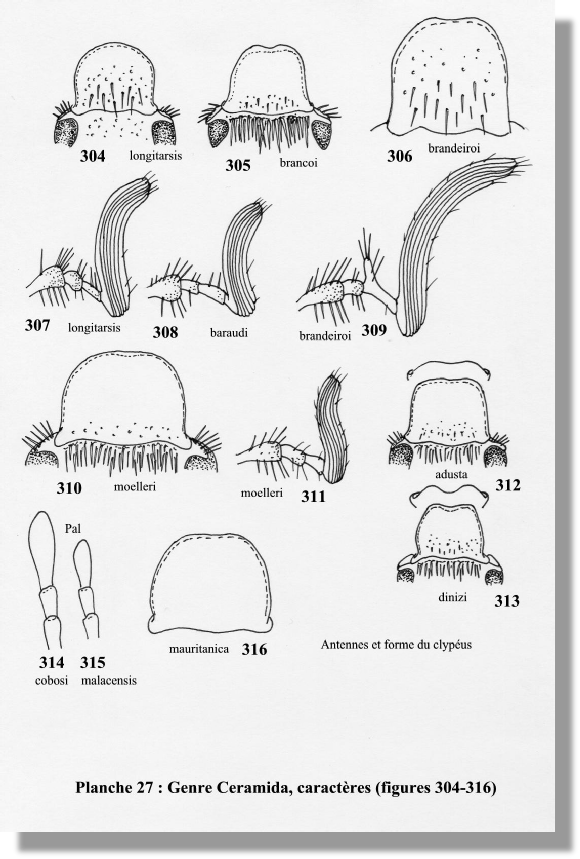
1.- Front glabre en arrière du sillon clypéo-frontal (fig. 304) 2
— Front avec une touffe de soies dense en arrière du sillon (fig. 305) 4
2.- Longueur du corps inférieure à 10,5 mm ; clypéus à côtés subparallèles en arrière (fig. 306) ; massue antennaire 2,5 fois plus longue que le funicule (fig. 309) ; Portugal brandeiroi
— Longueur du corps supérieure à 10,5 mm ; clypéus à côtés arrondis ; massue antennaire inférieure à 2 fois la longueur du funicule 3
3.- Massue antennaire 1,8 fois plus longue que le funicule (fig. 307) ; tégument du pronotum lisse et luisant ; Portugal longitarsis
— Massue antennaire 1,4 fois plus longue que le funicule (fig. 308) ; tégument du pronotum microponctué ; Portugal ; Espagne baraudi
4.- Ratio massue antennaire/longueur du corps plus petit que 0,120 5
— Ratio massue antennaire/longueur du corps plus grand que 0,120 6
5.- Espèce de grande taille (17,5 mm – 24 mm) ; massue antennaire plus longue que le funicule (fig. 311) ; Portugal ; Espagne moelleri
— Espèce de plus petite taille (12 – 16 mm) ; massue antennaire plus courte que le funicule ; Portugal ; Espagne bedeaui
6.- Ratio massue antennaire/longueur du corps plus grand que 0,145 7
— Ratio massue antennaire/longueur du corps entre 0,120 et 0,145 8
7.- Taille supérieure à 14 mm ; clypéus faiblement échancré au milieu (fig. 312) ; Espagne adusta
— Taille entre 10 et 12 mm ; clypéus fortement échancré au milieu (fig. 313) ; Portugal dinizi
8.- Dernier article du palpe maxillaire plus long que les deux précédents (fig. 314) ; clypéus glabre ; taille : 14 – 18 mm ; Espagne cobosi
— Dernier article du palpe maxillaire plus court que les deux précédents (fig. 315) 9
9.- Clypéus étroit, trapézoïdal, avec le bord antérieur ayant une forte échancrure médiane (fig. 305) ; Portugal brancoi
— Clypéus large, à côtés arrondis et bord antérieur peu échancré (fig. 316) 10
10.- Clypéus très concave, ponctué seulement vers sa base ; massue antennaire plus longue que le funicule ; ponctuation du pronotum très fine, peu visible ; Espagne ; Portugal ; Maroc malacensis
— Clypéus peu concave, entièrement ponctué ; massue antennaire pas plus longue que le funicule ; ponctuation du pronotum forte, bien visible ; Maroc ; Algérie mauritanica



Ceramida bedeaui Ceramida malacensis

Ceramida adusta (Kraatz) (12) (fig. 312)
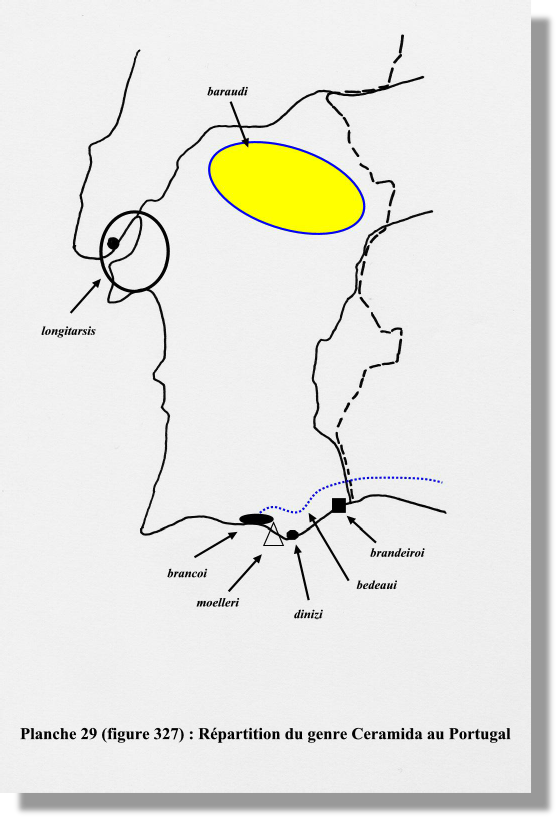
Distribution. – ESPAGNE : Cordoue, Séville, Huelva.
Biologie. – Le comportement de C. abderramani (= adusta) a été observé par López-Colón, 1993. Ces observations ont été effectuées du 5 au 12 novembre 1989 dans les provinces de Séville (Aznalcazar) et de Huelva (Hinojos). Le nombre d’exemplaires étudiés s’élève à 240 (dont seulement trois femelles). Activité diurne, entre 7 heures du matin et 13 heures de l’après-midi. Journées ensoleillées, sans pluie et sans vent. Les mâles déambulent sur le sol ou volent lourdement au ras du sol à la recherche des femelles. L’unique activité des mâles est la quête des femelles. Celles-ci sont en permanence enterrées à faible profondeur parmi le système radiculaire de la graminée Piptatherum miliaceum dont les racines nourrissent les larves. Les mâles vivent peu de jours. Ils s’enterrent pour la nuit à une profondeur de 1-5 cm. L’activité est principalement matinale. Les accouplements ont lieu durant les jours ensoleillés d’automne par une humidité de +80 % et une température de 15-18°C. Les larves de l’élatéride Stenagostus laufferi Reitter, 1904 sont prédatrices des larves de Ceramida. Les imagos moribonds et les cadavres sont dévorés par des ténébrionides du genre Pimelia F. L’espèce C. abderramani a pour les imagos une courte période annuelle d’apparition. Les mâles ne s’alimentent pas. Les femelles sont peu mobiles et se tiennent auprès de la plante nourricière des larves. La localisation de la femelle par le mâle s’effectue au moyen de phéromones émis et détectés par l’énorme massue antennaire du mâle. Le développement complet de la larve dure plus de deux ans.
Ceramida baraudi (Branco) (13) (fig. 308)
Ceramida bedeaui (Erichson) (14) (figs. 289-303)
Distribution. – ESPAGNE : Huelva, Cadiz, Malaga, Andalousie, Gibraltar.
PORTUGAL : Algarve : Ludo, Faro, Monte Gordo, Vila Real.
Biologie. – (D’après Rambur, 1843) : abondants autour de la baie de Cadiz, sur le sable des dunes, au mois de février. Accouplement le jour et lorsqu’il pleut. Femelle au sol à demi-enterrée, entourée par de nombreux mâles. Disparaissent lorsque les pluies cessent. (Serrano, 1985) : propre au littoral avec des forêts de pins, dans l’Algarve.
Ceramida brancoi (Baraud) (15) (fig. 305)
Distribution. – PORTUGAL : Algarve (IX,X, sous la pluie).
Ceramida brandeiroi (Flach) (16) (figs. 306, 309)
Distribution. – PORTUGAL : Algarve (Faro, Castro Marim).
Ceramida malacensis (Rambur) (20) (fig. 315)
Distribution. – ESPAGNE : malacensis : Andalousie, plage de Malaga (Rambur, fin de l’automne) ; mancinii : Séville, Cadiz ; luctuosa : Cadiz, Chiclana de la Frontera.
PORTUGAL : lusitanica : Ribatejo.
MAROC : tangeriana : Tanger, Larache.
Ceramida mauritanica (Rambur) (21) (fig. 316)
Distribution. – MAROC : Melilla. ALGERIE : Alger, Oran, Nemours.
Biologie. – Lucas (1849) : pris le soir, au printemps, après le coucher du soleil, le long des tiges de Asphodelus ramosus, aux environs d’Oran, dans les ravins.
Ceramida moelleri (Flach) (22) (figs. 310, 311, 317-326)
Distribution. – PORTUGAL : Algarve : Praia de Falésia, Aldeia das Açoteias.
ESPAGNE : Andalousie : Huelva.
Ceramida ohausi (Reitter) (23)
Distribution. – ESPAGNE ? (indiqué de Grèce par Ohaus, localité fausse selon Reitter qui la suppose d’Espagne).


Pour avoir la bibliographie complète ainsi que les synonymies nombreuses pour les espèces de ce genre, il faudra consulter le « Catalogue » que l’on pourra télécharger au format PDF sur ce site.

< Ceramida sp.